1. 文献 ¶
Hobson, H. M., & Bishop, D. V. M. (2017). The interpretation of mu suppression as an index of mirror neuron activity: Past, present and future. Royal Society Open Science, 4(3), 160662–. https://doi.org/10.1098/rsos.160662 论文原文
2.研究目的 ¶
本篇综述有三个研究目的,一是旨在回顾mu的过去,二是概述当代mu抑制研究中的问题(α问题;mu是否是MNS好的测量的问题),三是审视最近mu用于研究MNS在语言,社交过程和ASD中的参与,Mu抑制研究在很大程度上未能提供MNS在这些领域中作用的有力证据。(虽然传统上对mu抑制的评估是对简单的动作观察做出的响应,并且MNS的主要功能被认为是对动作的理解,但当代理论已经扩展,以假定MNS在语言,社会过程中的作用和ASD中的功能,这篇综述还将考虑mu抑制的研究是否可以阐明MNS的这些额外的假定功能。)
3.什么是mu节律 ¶
Mu是在典型的人类脑电图(EEG)中观察到的一种节律,通常定义为8-13 Hz频带,认为是由感觉运动区出现的。近年来,mu功率(power)的变化(即mu频带的强度)已被用作研究人体镜像神经元系统(MNS)的一种手段。 通常,检验这些变化的研究以mu抑制研究的形式进行,其中将mu的功率(power)在基线条件(预计不会激发镜像神经元活动)和实验条件之间进行比较。
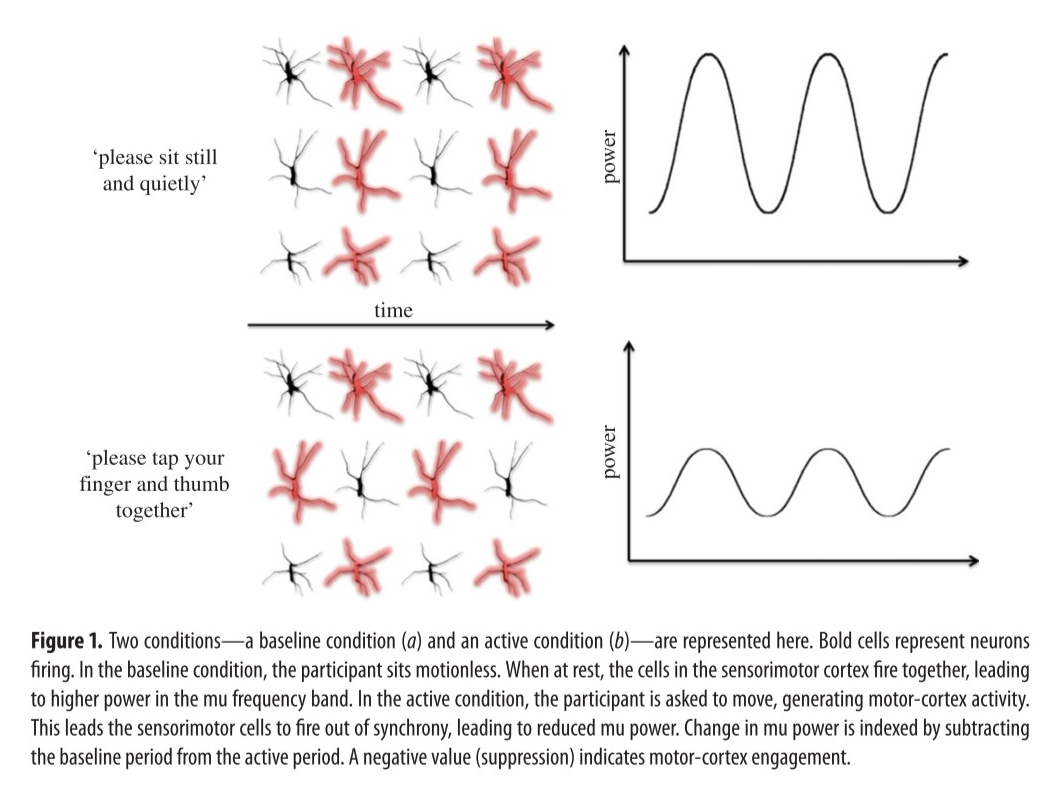
3.1 关于mu抑制的研究 ¶
Mu抑制研究的数量在最近十年已经增加了。最近的荟萃分析研究了1990年以来进行的85项研究(包括来自1707名被试的数据)。这仅包括检验典型被试的mu节律活动并使用具有动作观察条件或动作执行条件(或两者皆有)的实验范式的研究[1]。 在这85项研究中,有49项是自2010年以来进行的。Cuevas及其同事[2]还指出,mu抑制研究在婴儿中的应用有所增加。
4. Mu 抑制:简史 ¶
4.1 mu抑制的早期研究:“en arceau的节律” ¶
人类的μ节律研究已有很长的历史,可追溯到一些最早的EEG实验,早于发现镜像神经元。
人类mu 节律最初是由法国科学家Henri Gastaut [10,11]所描述的,由于它是波浪的拱形或小门形状,被称为rolandic wicket 节律或 rolandique en arceau节律。 早期的观察发现,这些波似乎是从rolandic区发出的,其速率为大约7-11 s(在当代文献中,mu频带(frequency band)通常定义为8-13 Hz)。
在1950年代,mu节律被认为是一种罕见的现象,甚至被认为是精神病理学的指征。 Schnell&Klass [12]仅在被试的2.9%中确定了mu。Gastaut等人[13,14]在500名健康的男性成年人中,发现仅有14%的人有mu,其发生被认为是与精神病性人格特征有关。在这里可以联想到自闭症谱系障碍(ASD)与mu异常有关的最新工作,并试图对该人群使用神经反馈。 参见§4.3。
正如Niedermeyer&Silva [17]所指出的,标准化帽的出现使得从脑电图C3和C4位(位于感觉运动皮层的中央位点)常规识别mu节律变得更加容易,并且用于分析EEG的新技术也证实了mu的发生比以前想象的更普遍。 Kuhlman [18]指出,在被试的脑电图中很少能以目视的方式识别出具有特征性门形状的mu活动(即通过肉眼观察的方式识别mu波),但脑电功率谱分析(power spectral analysis)在大约一半的样本中显示出这种活动。通过计算一致性,Schoppenhorst&Brauer [19]能够识别54位被试中60%的mu,比以前发现的比例高得多。他们还建议,不同的警惕程度的效果,可能会导致在某些被试中难以识别mu。
很明显,mu不是异常,特别要说是,它也不是病理现象, 关于mu节律可能与什么相关的新理论出现了。一种著名的理论是,mu代表感觉运动皮层的静止活动,抑制这种节律反映出大脑的这些区域变得活跃[18,19]。确实,在Gastaut的早期研究中,我们认识到,被试自己的运动会阻止mu的节律,进一步的实验发现mu不仅会被自发运动阻止,还会受到被动运动,反射活动和运动命令的阻碍[20]。
事件相关的不同步或同步(ERD / ERS)描述了给定功率带(power band)相对于基线的减小或增大。在很大程度上,ERD和ERS被认为分别反映了皮质激活和空闲(idling)。 Pfurtscheller&Lopes da Silva [21]记录了与α/μ(8-13 Hz)和β(13-35 Hz)频段活动有关的ERD / ERS使用的基本原理。
研究人员使用ERD / S证明了mu对人们自身运动的反应,并建议可能存在不同类型的mu节律,也许对应于运动皮层的不同区域[22]。然而,正是mu对其他形式的刺激产生了响应,使得在过去十年中产生了大量研究。
4.2 Mu的抑制和镜像神经元的发现 ¶
除了对被试自己的运动做出反应之外,通过观察其他人的运动也会抑制mu。随着科学家开始研究脑电图对电影放映的反应,出现了在观察运动过程中mu抑制的报道。
在1980年代,一组意大利神经科学家发现了猕猴大脑中的细胞,它们在动物执行动作时以及观察到另一动物正在执行的动作时均会触发[4,5]。 这些细胞随后被命名为镜像神经元,观察运动时感觉运动皮层被激活的现象演变成对镜像神经元的动作理论的理论[3]。在猕猴中发现镜像神经元之后,mu抑制现象有了新的解释。可以说,μ频带可辩证的显示出与镜像神经元相似的响应特性。将mu和镜像神经元相提并论(Parallels were drawn between mu and mirror neurons),并且mu活动性的降低被认为是镜像神经元活动的标志[23,24]。
最初在猴子身上进行的实验表明,镜像神经元的活动与目标定向动作特别相关;这些经典研究使用的刺激显示手与物体相互作用。在动物文献中,不指向物体的等价的运动不会引起镜像神经元活动[4]。对人类被试的Mu抑制研究发现,在看到其他人的手精确的抓握(即可以在物体上使用的抓握)而不是中立的、非抓握位置时,会产生更强的mu抑制,并且物体相互作用比没有物体相互作用的条件产生更大的mu抑制[23,24]。已经提出,该物体效应(object effect)是μ抑制与人类镜像神经元活动有关的证据。然而,可以说,对动物记录工作的严格解释将表明,在观看与物体无关的动作时,根本不应该发生mu抑制。相反,一些作者推测,MNS对非物体定向行为的反应是人类镜像神经元的显著特征,并且与其他灵长类动物的这种差异可能代表了与我们共同祖先的背离。进一步提出,这种发展可能在语言的发展中发挥了重要作用[25]。但是,更乏味的解释是mu抑制可能正在测量MNS下游区域的活动,,而不是镜像神经元区域本身[24]。((Muthukumaraswamy, S., Johnson, B., & McNair, N. (2004) Mu rhythm modulation during observation of an object-directed grasp. Cogn. Brain Res. 19,195–201. http://doi.org/10.1016/j.cogbrainres.2003.12.001)
这些开创性研究表明可以利用mu抑制来研究人类MNS,因此进行了更多的动作观察实验。Fox等人[1]通过荟萃分析对此问题进行评估得出的结论是,mu抑制可以用作人类镜像神经元活动的代理。
然而,其他人争辩说mu抑制反映了镜像神经元的活动。如:Coll等[26]报告说,μ抑制是感觉镜像而不是运动镜像的指标,这一发现破坏了认为镜像神经元代表的动作和感知之间的重要联系。
此外,Fox等人的结论[1]被Hobson & Bishop[27]挑战。本篇作者认为,mu抑制的实验方法通常会与非镜像过程混淆。在本篇作者自己的结果中,在动作观察过程中的mu抑制并非特定于生物运动,或者不一定特定于位于感觉运动带(sensorimotor strip)上方的中央电极。因此,mu抑制作为人类MNS测量的有效性是当前争论的话题。 (Hobson, H.M., & Bishop, D.V.M. 2016 Mu suppression—a good measure of the human mirror neuron system? Cortex 82, 290–310.http://doi.org/10.1016/j.cortex.2016.03.019) (查看以上文献,查证
(1).是否mu抑制特定于生物运动,非生物运动指的是什么,是否特定于中央电极,是否只是特定于运动观察情况下; ¶
(2).如果不特定于运动观察,是否在运动想象条件下,不特定于中央电极,非生物运动,也有mu波抑制的产生;非身体物体的旋转,是否可以算作非生物的运动; ¶
(3).如果mu波不特定于中央电极,也可能在枕电极,是否无法判断位于枕叶的是α波还是mu波。) ¶
5. mu抑制是镜像神经元系统的好的测量吗? ¶
5.1 mu抑制研究的科学质量 ¶
由于mu抑制已在认知神经科学中广泛用于推断高级社会过程和临床疾病中镜像系统的作用,因此mu抑制是否能很好地反映镜像神经元活动的问题成为重要的问题。最近的一项荟萃分析得出的结论是,尽管mu抑制提供了研究MNS参与的有效手段,但在文献中仍存在一些局限性[1]。这些问题包括相对于mu抑制领域相对特定的问题,包括以下事实:很少有研究报告中心电极以外位置的功率(power)变化,因此无法确定影响不是由其他位置的功率(power)变化驱动的。一个相关的问题是,如果条件之间存在注意差异,则可能会在另一个功率(power)信号,α中产生广泛的变化,从而可以模仿mu抑制。我们附和(echo)这些建议,但也注意到mu抑制文献中的其他一些问题,其中一些问题也适用于神经影像学和心理学的更广泛领域。我们在讨论针对mu抑制研究的一些设计问题之前,首先考虑这些更广泛的观点。
5.1.1 小样本问题 ¶
首先,mu抑制研究通常遭受小样本量和因此低的统计能力。在使用自闭症等临床组的研究中,这是可以理解的,因为这些组可能难以招募,并且对EEG程序的耐受性可能较差。但是,即使对于非临床样本,也习惯使用20个或者不超过20个的样本。容易领会(appreciate)的是,小样本量会降低检测到真实效果的可能性。但是,通常假定如果一个效应已经被发现了,即使样本量很小,这个效应也是真的。然而,这种假设是不准确的-缺乏功率(power)也意味着,一个显著的效应可能并不会反映真实效应[28]。在神经科学领域,低功率(power)现象很普遍,在随后的研究中增加了假阳性效应的风险,高估效应大小和再现问题(problems reproducing effects)的风险[28]。
一项给定研究所需的被试数量取决于许多因素,包括分析设计,条件数量,预期效应大小,测量之间的相关性等等。因此,mu抑制研究并没有给定被试数量。然而,作为一个粗略的指导,具有两因子的重复测量设计每个都包含双向ANOVA分析的两个水平,则需要40名被试,才足够来检测具有90%功率(power)的中型主效应1。检测交互作用,需要47名被试。
5.1.2 频率段问题 ¶
其次,mu抑制是一种具有大量的分析灵活性的现象,这是重现性差的另一个已知风险因素[31]。例如,mu抑制研究在哪个频段被认为是“ mu”上不同。频段不是独特的类别,而是EEG文献中提出的灵活范围,这意味着mu抑制论文可以采用彼此略有不同的频段。 “ mu 频带”在先前的实验中已定义为:8–12Hz(例如[32]),8–13Hz(例如[33,34]),8–15Hz(例如[35]),8–16Hz [36] ],10–14Hz [37]或分成上,下活动频段(例如[38,39])。确实,尽管许多mu抑制实验将mu定义为alpha波段(8–13 Hz)活动,但mu波实际上被认为是由来自两个频率的贡献组成的,包括alpha和beta(13–30 Hz),并且在大约10和大约20 Hz具有典型的峰。
一些研究表明,β波段而不是α波段的活动可能是MNS参与的更好指标(但是,参见[27])。因此,一些研究基于α-mu和β-mu可能具有不同的响应模式,或同时检验了α和β的活动,以检验较高和较低的μ频带。
其他研究人员认为,可能必须根据每个人不同情况计算出正确的频带,类似于磁共振成像中功能定义的位置(functionally defined sites)。这可能尤其重要,因为mu节律被认为是神经反馈的目标,并且已经提出了计算个人频带的方法 [21]。尽管可能确实有理论上的逻辑依据可用于分割 mu节律,或选择更高或更低,更窄或更宽的频段进行检验,但如果这些决定是基于要分析的相同EEG数据,则存在疑问。这为研究人员提供了一个选择频段的余地,从而可以提供最佳结果以适应他们的假设,包括在分析中引入循环(circularity)[40]。
5.1.3 方差分析的问题 ¶
与分析灵活性问题相关的是,计算大量相关性或运行方差分析的研究,而没有对多重测试进行适当的校正[41]。这些研究在设计上可以说是探索性的,因此需要把它们看作是探索性的。尽管ANOVA有效地校正了给定因子内的水平数量,但它们不会自动校正因子数量或因子之间潜在相互作用的数量。例如,三向方差分析用于测试三个主效应,三个双向交互作用和一个三向交互作用。在这种方差分析中,根本没有发现明显效应的可能性(如果将alpha设置为0.05)为0.95的七次方 = 0.70。因此,获得重大的主效应或相互作用的可能性为30%。对于四向方差分析,这种可能性增加到54%。方差分析不仅在mu抑制文献中是有问题的,而且在更广泛的EEG领域和行为科学中也是有问题的[42](另请参阅Bishop的博客文章[43],以讨论与EEG相关的这些问题),如Luck&Gaspelin所述 [44],即使在最近发表的脑电图实验中,这些问题仍然很普遍。这些问题的示例特别是在第4.2.1节。
5.2 α的问题 ¶
在mu抑制研究中,可能难以确保由感觉运动区域引起的8-13Hz频带的变化,是由与镜像神经元相关的过程引起的,而不是由脑中的其他区域,或其他同时发生的过程引起。如果在动作观察过程中发生与镜像神经元相关的过程,则需要在无数其他认知和知觉过程的背景下进行检测,这些过程可能会掩盖或混淆它们的检测。确实,可以在许多区域观察到该频段的活动,通常称为alpha频段活动,并且其变化已牵涉到许多过程中[45]。
所谓的可以将“ mu”与枕骨“ alpha”区分开来的是地形和反应性/响应率/敏感度(responsivity)——尽管alpha在枕骨皮层中最为突出,并且对视觉刺激和注意力的变化会有所反应,mu仅限于感觉运动区域的电极并对被试自己的动作作出反应。当然,在动作观察过程中,很有可能感觉运动相关的mu抑制和注意力相关的alpha抑制都将独立发生;
(自注:ter Horst, A.C., van Lier, R., & Steenbergen, B(2013)中,认为视觉想象引发了枕区的mu抑制,但是很有可能这是由于注意相关的α抑制导致的)
发现重要的枕骨α抑制并不排除MNS的参与已经发生了,这种可能性。尽管如此,研究人员有责任解开来自其他与注意力和知觉有关的认知过程的镜像神经元活动之谜。
5.2.1 研究表明mu与α的分隔是不充分的 ¶
也许最能阐明这一点的研究是那些,考虑了mu抑制与其他旨在测量MNS的措施之间的相关程度如何。此类研究包括同时进行脑电图和功能磁共振成像(fMRI)记录的那些研究,以研究这两种措施是否一致,以及mu抑制是否可以作为研究MNS的更便宜,更容易获得的方式[46–49]。
很明显,该结果是积极的——被认为是人类MNS(包括下顶叶,背侧前运动和初级躯体感觉皮层)的一部分的大脑区域的BOLD反应与mu抑制相关。有趣的是,尽管先前的作者推测mu抑制最可能是由Broca区产生的,这是围绕MNS和语言的理论的一个关键论点(请参见§4.1),Arnstein等人的发现[46]不支持该概念。然而,尽管存在这些相关性,但作者提醒说,他们的结果还表明mu抑制也可能反映了其他网络的活动,包括涉及视觉运动过程的区域(该区域并非是MNS的一部分[47,49])。
(自注:mu抑制对涉及视觉运动过程的区域活动的反应,(1)可能是对视觉心理想象的反应,参见ter Horst, A.C., van Lier, R., & Steenbergen, B(2013);(2)也可能是由于警觉度,注意程度不同造成的影响) (因为:alpha在枕骨皮层中最为突出,并且对视觉刺激和注意力的变化会有所反应,mu仅限于感觉运动区域的电极并对被试自己的动作作出反应。)
MNS活动的另一个一般认定的指标是经颅磁刺激(TMS)诱导的运动诱发电位(MEPs)。 Lepage等 [50]结合脑电图和TMS来研究这两种措施之间的关系。与以前的研究一样, MEP幅度显著增加,在动作观察,想象和执行过程中,对mu的抑制也得到了显示,但这些措施之间没有关联。 mu抑制有效性的其他证据来自脑磁图(MEG)研究。 MEG在空间上对源进行定位的能力被认为优于EEG。在MEG mu抑制研究中,通过观察和执行显著调节的区域是感觉运动皮层,但在枕骨区域也发现了效应[51,52]。
mu与alpha之间的分隔不充分可能潜在得意味着报告的对某些刺激的mu波的反应实际上是阿尔法的变化,反映了条件之间注意力的差异。例如,Aleksandrov&Tugin [53]在一系列控制条件下测量了mu抑制,包括精神计算周期(periods of mental counting)或观察非生物物体的运动,这些条件下我们不会预测镜像神经元会变得活跃。然而,在这些条件下对mu的抑制并不明显小于被试观察到人类行为这种条件下的mu抑制。此外,对注意要求最高的任务似乎产生了最强的mu抑制作用,这表明心智努力(mental efforts)可能会混淆/搞乱(confound)mu研究。
同样,Perry&Bentin [54]指出,在他们的mu抑制研究中,条件之间的显著差异可能是由于注意需求的差异,而不是镜像神经元活动的差异所致,这是在找到枕骨和中央电极的类似的8-13HZ 功率(power)的变化模式后说明的。alpha的经典标记之一是睁眼的阻塞(blocking);在一项研究中,睁眼对mu抑制会有最大效果,大于任何实验条件[38]。
有充分的文献记载,阿尔法活动与机敏性和认知努力(cognitive effort)有关,并且一些最早的报告指出,mu也是如此。
Chatrian等人[20,55]报告说,mu波取决于警觉度的变化,而心算(mental arithmetic)[20]或问题解决[56]会抑制mu节律。 Schoppenhorst&Brauer [19]说:“虽然在视觉上评估常规脑电图时,我们注意到mu波非常不稳定。 仔细观察我们发现,这种能力可以归因于轻微警觉度的变化,通常很难从阿尔法节律的变化中辩别出来”(第25页)。 后来:‘由于它们对警戒程度的依赖性,它们非常不稳定,并且可以通过增加或减少警觉来抑制它们。”(第31页)。
Perry&Bentin [54]警示说,“mu抑制报告不仅应始终包括在中央电极的实验效应,而且还应包括枕骨区域,以帮助充分了解所研究的现象”(第1054页)。 然而,考虑到枕电极活动的μ抑制研究却有混合发现。 Lepage等[50]在同时进行的TMS-EEG研究中,从枕叶(Oz)上的电极进入活动进行分析,发现在被试观察到动作或想象自己执行动作的条件下,此部位的8-13Hz功率(power)确实受到抑制[50]。
然而,针对自闭症被试异常mu反应的mu抑制研究报告说,除了C3,Cz和C4以外,没有其他电极显示出一致的抑制模式,这表明由于枕骨α变化引起的混淆,他们的结果模式不可能出现[57-60]。
5.2.2 基线问题 ¶
总结:基线期的选择会对是否能观察到mu抑制产生影响:如果基线期被试的α波受到注意的影响,可能会测不到显著的ERD.
与α和注意力相关的问题有关,在mu抑制实验中选择基线技术可能对是否观察到mu抑制起重要作用。正如Pfurtscheller&Lopes da Silva [21]所指出的那样,“ ERD是以相对于参考间隔的功率(power)百分比来衡量的,因此,ERD取决于该间隔内的节律活动的量”(第1847页)。不活动的基线(被试没有受到刺激并且被要求安静坐着的位置)可能会在基线期间使alpha功率(power)膨胀(inflate)。
然后,当从该时期减去实验条件时,观察到8-13Hz频段显著下降的可能性增加,这被认为是mu抑制,因此运动皮层或MNS参与。潜在地,基线的不同选择可能会导致mu抑制文献中不一致的发现。而Fox等人的荟萃分析[1]提示:基线不影响是否观察到mu抑制,某些基线技术已突出显示了问题 [27]。
6. 超越动作理解的Mu抑制 ¶
在阐明了mu抑制的潜在缺点之后,我们现在探索使用该技术探索MNS在许多不同功能中的作用的研究结果。虽然最初MNS被提议主要用作行动理解的基础,但是MNS可能扮演的潜在角色很快就扩大了。语言,心理理论和同理心都被认为扎根于MNS。如果mu抑制是对人类MNS的充分度量,并且如果MNS参与了这些过程,那么人们会期望在涉及语言,心理理论和同理心的任务中看到mu抑制。在这里,从mu抑制研究中发现镜神经元参与这些过程的证据。
我们从(Psych Info)中搜索“ mu抑制”,“语言”,“社交”或“自闭症”来获取消息来源。还从阅读的已发表论文的参考文献中获得了其他相关研究。其中包括以英语发表的,使用人类被试的文章(已有一些发表的实验,使用类似的EEG技术检查非人类灵长类动物的镜像神经元功能)。这篇综述的重点是成年人,但是对于使用自闭症被试的研究,这些样本还包含儿童。在评估有关mu抑制和MNS的论文时,我们发现所有论文都研究了一个较低的频带(约8–13 Hz),其中一些还考虑了beta频带(13–30 Hz),但没有一篇论文专门讨论Beta。我们寻求过去十年间(自2007年以来),mu被重新概念化为镜像神经元活动的代理之后的研究。因此,本综述捕获了由镜像神经元相关理论所激发的实验中mu抑制使用的最新趋势。
6.1 语音和语言任务中的Mu抑制 ¶
6.1.1 语言任务中mu抑制的概述 ¶
语言的mu抑制研究主要集中于语音感知和产生,而不是语义理解。 这类研究得出的结论颇为复杂。许多人至少在某些任务中报告了抑制,但是在某些情况下,方法论问题降低了对发现的信心。一些研究者还主张对交流障碍中的mu抑制进行研究,尽管目前还没有关于语言障碍人群中mu抑制的研究[78]。
考虑到语音感知中mu抑制的证据(与语言理解相反),在被动听之外,需要额外处理的任务似乎更容易出现抑制。两种理论可以解释这一点。首先,仅在语音处理足够需求时,才出现运动区域的募集(recruitment of motor)。运动区域被有效地“引入”作为任务的附加资源,这表明基于运动皮质的语音感知不是处理语音的唯一方法,而是一种附加的支持,而不是言语感知的核心过程(见[74,79])。这将表现出:MNS-语言理论的弱得多的版本。第二种理论,或者说更简化的理论是,当语音感知(或任何其他一种)任务变得足够困难时,由于注意力的影响,可以看到alpha频段的抑制。 Alpha频段的活动遍布整个头部,而不仅仅是在感觉运动皮层,并且任务投入和精神活动的变化,会导致8-13Hz频段的抑制,这可能与运动皮质的活动无关。
最后,在同一实验中还缺乏研究在知觉和产生中检测mu抑制的研究,鉴于言语知觉的运动理论的难题/关键,是支持知觉和产生的基质(substrate)之间的重叠,因此这项测试显得至关重要。
总而言之,在多大程度上语音和语言任务导致mu抑制独立于其他潜在的混杂因素(confounds)存在争议,如果MNS在语音和语言过程中起关键作用,那mu对语音刺激的反应在多大程度上符合预期。如果可以在预先注册的设计(pre-registered design)中复制某些关键研究,则可以对发现有更大的信心,在该设计中,在检查数据之前要确定假设,方法和分析方法,以防止追溯假设的拟合(retrospective hypothesis fitting)。这些理论做出了明确的,可检验的预测,并且似乎非常适合这种证据收集方法(有关复制和预先注册(pre-registration)的良好指南,参见[80])。
6.2 社交过程中的Mu抑制 ¶
包括在移情,心理理论,生物运动中的mu抑制。
6.2.1 生物运动 ¶
一些研究已经考虑了mu对生物点光源展示的反应。这些展示是通过用光标记运动物体的肢体运动而创建的图像序列。这些刺激提供了一个解决方案,可以提供匹配良好的刺激来研究mu响应-社会与非社会刺激通常在许多基本的感知因素上有所不同,而点光源显示则可以更严格地控制这些变量。
对这些显示的Mu抑制已被用来证明镜像神经元参与了生物运动的处理。在对20位被试的研究中,Ulloa和Pineda [93]发现了对生物点光源展示的显著mu抑制,但对扰动运动(scrambled motion display)展示却没有。他们争辩说,其效果不是由于注意力的不同所致,因为连续执行任务的执行情况在这些条件下没有差异——但是,未报告中央电极以外区域的结果。
确实,其他检查mu抑制点光展示的作者提出了告诫,从枕骨alpha和不同条件下的注意差异可能引起潜在的混淆效应。 佩里等[88]研究了被试识别表现在点光源展示上,所代表的不同维度的能力(所描绘的情感,模型的性别(gender of the model),非生物点光源显示的行走方向和滚动方向)。与非生物运动条件下的滚动方向相比,被试对展示中所代表的某些社会维度(情绪,性别,意图)做出决策的速度较慢,有时准确性较低,非生物运动条件下的滚动方向,表明这些任务与任务难度不匹配。此外,在对24位被试的EEG数据进行分析时,他们报告了枕叶区域的结果,这些结果显示在整个条件下均具有显著的α抑制作用,并且其抑制模式与中心部位相似。作者建议,生物点光源显示可能会引起更多关注,因为这些因素具有更高的生态价值(例如,需要知道某人是朝着你走还是从你这里走开)。
6.2.2 概述(待修改) ¶
迄今为止,将mu抑制与社会过程相联系的发现是多种多样的。试图使用mu抑制作为MNS的个体差异测量的研究似乎特别有问题。几乎没有观察到强烈的相关性(几乎没有一个在多次比较的校正后仍然存在),并且尚不清楚其他与镜像神经元无关的理论,是否可以解释其中的一些发现。当然,许多研究并未充分考虑设计中阿尔法或注意效应的潜在混杂因素([88]是一个明显的例外,清楚地表明了考虑此类问题的重要性)。另一个考虑因素是被试自己的动作是否会混淆效果。被试产生自己运动显然会导致运动皮层的参与和mu抑制。因此,如果条件能够改变被试执行的动作量,则这些差异可能会混淆mu抑制研究的结果。指示被试不要移动可能并不总是足够的,因为有证据表明个人可能在没有意识的情况下模仿他人。自动模仿是一种现象,个体在不知不觉中模仿了自己在他人身上感知到的动作或姿势。的确,由于有证据表明,对同情心得分高的被试比对低心智被试的模仿更多,自动模仿可以可行地调解已报道的共情mu抑制之间的关系[94]。在整个领域,许多mu抑制研究的确记录了被试的肌电图,以放弃被试移动的试验,但这可能是mu抑制和共情研究的一个特殊问题,因为这些论文很少讨论自动模仿。总体而言,在这些研究中似乎没有必要诉诸于镜像神经元理论的解释-替代解释(account—alternative accounts),包括社交刺激与自动模仿效应之间的注意差异,也可以解释这些结果。